Биология. 11 класс
§ 16. Клеточный цикл. Репликация ДНК
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Процесс репликации ДНК
Информация, записанная в ДНК, должна быть не только реализована в процессе развития клеток и организмов, но и в полном объеме передана следующему поколению. С этой целью перед делением клетки в ней осуществляется процесс репликации, т.е. удвоения количества ДНК.
Информация о механизме репликации содержится в самой ДНК: одни гены кодируют ферменты, синтезирующие предшественники ДНК — нуклеотиды, другие — ферменты, обеспечивающие соединение активированных нуклеотидов в единую цепочку. Механизм репликации был впервые постулирован Дж. Уотсоном и Ф. Криком, которые отмечали, что комплементарность цепей ДНК наводит на мысль, что эта молекула может удваивать саму себя. Они предположили, что для удвоения необходим разрыв водородных связей и расхождение цепей, каждая из которых играет роль матрицы при синтезе комплементарной цепи. В результате одного акта удвоения образуются две двунитиевые молекулы ДНК, в каждой из которых имеется одна материнская нить и одна новая (см. рис.).

Полуконсервативная репликация ДНК
Механизм получил название полуконсервативной репликации. Позже матричная природа и постулированный принцип репликации ДНК были подтверждены многочисленными экспериментальными данными.
Репликация ДНК начинается в специфических точках хромосомы — сайтах инициации репликации (origin). Процесс репликации обслуживается большим количеством ферментов. Наиболее полно изучен аппарат репликации бактериальной ДНК, особенно E. coli. Функцию расплетания молекулы ДНК у прокариот выполняют специфические ферменты геликазы, которые используют для работы энергию гидролиза АТФ до АДФ. Они часто функционируют в составе белкового комплекса, осуществляющего перемещение вилки и репликацию расплетенных нитей. Удерживают нити ДНК от воссоединения другие специфические белки, связывающиеся с одноцепочечными участками. Эти участки, разошедшиеся в разные стороны, образуют характерную структуру — репликативную вилку (вилку Кернса). Это — та часть молекулы ДНК, в которой в данный момент осуществляется синтез новой цепи. В продвижении вилки большую роль играет белок гираза, относящийся к разряду топологических изомераз. Он обнаружен только у бактерий. Гираза — это релаксирующий фермент, который, производя двунитиевые разрывы, снимает положительные (перед вилкой) и способствует образованию отрицательных (сзади вилки) супервитков в релаксированной ДНК.
Каждая цепь материнской ДНК служит матрицей для синтеза дочерних молекул. На одной цепи синтез осуществляется непрерывно в направлении от 5′ к 3′ концу. Эта цепь называется лидирующей. Вторая цепь с противоположной направленностью, называемая отстающей, синтезируется в виде отдельных фрагментов, которые затем сшиваются лигазами в непрерывную молекулу. Фрагменты названы по имени американского ученого Р. Оказаки, впервые постулировавшего такой способ синтеза ДНК, фрагментами Оказаки. В ходе синтеза репликативная вилка перемещается вдоль матрицы, и при этом новые участки ДНК последовательно расплетаются до тех пор, пока вилка не дойдет до точки окончания синтеза (точка терминации).
Схема репликации ДНК у Escherihia coli

Синтез новой цепи ДНК требует затравки в виде небольшого фрагмента РНК, т.к. ведущий его фермент ДНК-полимераза для работы нуждается в свободной 3’OH группе. У прокариот обнаружены три разных ДНК-полимеразы с аналогичными функциями, обозначаемые как polI, polII и polIII. Наиболее полно изучена ДНК-полимераза I. Она представляет собой одиночный полипептид с мультифункциональной активностью (полимеразной, 3′ → 5′ экзонуклеазной и 5′ → 3′ экзонуклеазной). Синтез затравки (праймера) осуществляет фермент праймаза, который иногда входит в состав комплекса — праймосомы из 15-20 белков, активирующих матрицу. Затравка состоит из 10-60 рибонуклеотидов. После того как ключевой фермент синтеза ДНК у E. coli — polIII — присоединяет к затравке первые дезоксирибонуклеотиды, она удаляется с помощью polI, обладающей 3′ → 5′ экзонуклеазной активностью, т.е. способностью отщеплять концевые нуклеотиды с 3′-конца цепи. Затравка синтезируется также и в отстающей цепи в начале каждого фрагмента Оказаки. Ее отщепление, а также удлинение фрагментов, синтезированных polIII, осуществляет polI. Роль polII в репликации ДНК E. coli до сих пор не совсем ясна.
Схема разных механизмов репликации ДНК у бактерий, эукариот и бактериофагов

При репликации ДНК эукариот процесс репликации осложняется присутствием в составе хромосом белков. Чтобы расплести ДНК, необходимо разрушить сильно конденсированный комплекс ДНК и гистонов, а после репликации вновь осуществить компактизацию дочерних молекул. Раскручивание ДНК вызывает суперспирализацию участков, расположенных рядом с репликационной вилкой. Для снятия возникающего напряжения и свободного продвижения вилки здесь работают специфические ферменты релаксации — топоизомеразы. В различных организмах идентифицированы два типа топоизомераз: I и II типов. Они изменяют степень сверхспирализации и тип сверхспирали, производя разрывы в одной (топоизомеразы I типа) или обеих цепях ДНК (топоизомеразы II типа), и устраняют риск спутывания нитей ДНК.
Репликация бактериальной ДНК является двунаправленным процессом с одним сайтом инициации. В отличие от этого хромосома эукариот состоит из отдельных участков репликации — репликонов и имеет много сайтов инициации. Репликоны могут реплицироваться в разное время и с разной скоростью. Скорость репликации ДНК в эукариотических клетках значительно ниже, чем в прокариотических. У E. coli скорость приблизительно равна 1500 п.н. в секунду, у эукариот — 10-100 п.н. в секунду. Двуцепочечные кольцевые ДНК некоторых вирусов реплицируются по типу катящегося кольца. В этом случае одна цепь ДНК надрезается в одном месте специфическим ферментом и к образовавшемуся свободному 3’ОН-концу с помощью фермента polIII начинают присоединяться нуклеотиды. Матрицей служит внутренняя кольцевая молекула. Надрезанная цепь при этом вытесняется, а затем удваивается по типу отстающей цепи E. coli с образованием фрагментов, которые сшиваются лигазами.
Читайте также другие статьи темы 6 «Молекулярные основы наследственности»:
Перейти к чтению других тем книги «Генетика и селекция. Теория. Задания. Ответы»:
Репликация ДНК: учебное пособие

Учебное пособие соответствует государственному образовательному стандарту дисциплин «Экология» и «Физико-химические основы цитологии» подготовки бакалавров по направлению 140400 «Техническая физика». В пособии описываются проблемы репликации ДНК. Излагаются современные представления о строении хромосом, координации в течение клеточного цикла процессов ДНК-метаболизма, а также описываются участвующие в этих процессах белки и рассматриваются механизмы, отвечающие за сохранение генетической стабильности организмов. Предназначено для студентов дневной, очно-заочной и заочной форм обучения, изучающих дисциплины «Экология» и «Физико-химические основы цитологии» в рамках подготовки бакалавров по направлению 140400 «Техническая физика».
Оглавление
Приведённый ознакомительный фрагмент книги Репликация ДНК: учебное пособие предоставлен нашим книжным партнёром — компанией ЛитРес.
Глава 2. Начало репликации
Репликация ДНК начинается не в любой случайной точке молекулы, а в специфических местах, называемых точками начала репликации или олриджинами. Процесс копирования продолжается через образование репликативных вилок в одном или обоих направлениях до тех пор, пока ДНК полностью не удвоится. В замкнутых кольцевых молекулах ДНК новосинтезированные цепи ковалентно соединяются в местах встречи увеличивающихся в размере репликативных вилок или в том месте, где единственная вилка возвращается к точке начала репликации. Дочерние молекулы, как правило, расходятся еще до начала нового раунда репликации.
Такие различающиеся по размеру геномы, как геном вируса SV40 (5,2тпн), бактериофага λ (48,5тпн) и Е. соli (4-10 3 тпн), воспроизводятся в результате одного инициирующего события, происходящего в определенной точке.

Рис. 2. Возможное движение репликативной вилки.
У про — и эукариот можно встретить различные вариации на эту тему. Так, каждая из цепей родительской спирали митохондриальной ДНК животных (15тпн) имеет свою точку начала репликации. Синтез комплементарной цепи некоторых небольших однонитевых фаговых геномов начинается вблизи одной специфической последовательности, а репликация полученного дуплекса может инициироваться совсем в другой точке. Репликация линейных двунитевых ДНК также инициируется в особых сайтах. Например, ДНК бактериофага Т7 (40тпн) реплицируется в двух противоположных направлениях к разным концам молекулы, начиная от одной точки, а каждая из двух цепей ДНК аденовируса человека (30–38 тпн) реплицируется последовательно всегда от З’-конца.
Для геномов эукариотических клеток характерно наличие множественных точек начала репликации, разбросанных по хромосоме на расстоянии около 20тпн. После инициации репликация продолжается в двух направлениях от каждой точки до тех пор, пока репликативные вилки двух соседних точек начала репликации не сольются. Полноразмерные ДНК каждой дочерней хромосомы получаются путем соединения более коротких, независимо инициированных новосинтезированных нитей.
2.1. Понятие о репликоне и ориджине репликации
Участок ДНК, на котором синтезируется отдельный фрагмент лидирующей нити, называется репликоном. У многих прокариот их геном содержит только одну точку инициации репликации, то есть у них в ДНК только один репликон. Эукариотические геномы полирепликонны.
Место начала репликона, в котором происходит инициация репликации, носит название ориджина репликации. Именно ориджин распознается специальными белковыми комплексами и на нем начинается формирование вилки репликации.
В некоторых случаях место начала репликации имеет такую нуклеотидную последовательность, что дуплекс принимает необычную конфигурацию, которую распознают белки, участвующие в инициации. Природа взаимодействия между точкой начала репликации и белками и механизм инициации в целом исследованы недостаточно, однако можно сказать, что, по-видимому, они в разных случаях различны.
2.2. Ориждин репликации E.coIi oriC
Наиболее подробно изучены ориджины у Е. соli и Bacillus subtilis. Область начала репликации хромосомы, оriС (origin of chromosome), включает в себя участки со специфическими последовательностями, так называемыми ДНК-боксами, и расположенными между ними короткими последовательностями. ДНК-боксы со специфическим «мотивом» нуклеотидов, преимущественно в 9пн, перемежаются фрагментами в 12-1Зпн с высоким содержанием АТ. Сами девятичленные последовательности могут располагаться как в прямом, так и в инвертированном положении по отношению друг к другу. Например, у В. subtilis имеется один фрагмент ТТАТССАСА и два других девятичленных бокса, ориентированных в противоположном направлении, с заменой одной из пар нуклеотидов. Всего у В. subtilis на оriС расположено 15 ДНК-боксов. Область оriС очень консервативна: ДНК-боксы сходного состава имеются в соответствующем месте хромосомы у других бактерий (только у Mycoplаsma genitalium, несмотря на наличие общих для всех бактерий ферментов репликации, ДНК-боксов найдено не было). Сами ДНК-боксы не кодируют белок или РНК, хотя между ними располагаются отдельные гены. Продукты этих генов также большей частью вовлечены в «обслуживание» процесса репликации ДНК.
Порядок расположения ДНК-боксов, промежуточных областей и их количество позволяют думать, что эволюционная дивергенция oriС шла главным образом за счет дупликаций и трипликаций. Схема абстрактного «минимального ориджина» прокариот представлена на рис. 3.

Рис. 3. Организация минимального ориджина прокариот
Схема минимального ориджина прокариот.
2.3. Ориджины других организмов
Коровая часть ориджина репликации у вируса SV40 состоит из элемента опознания (ORE — origin recognition element), необходимого для связывания особого белка Т-антигена (Т-аg), элемента для связывания белка, расплетающего ДНК (DUE — DNA unwinding element), и элемента, обогащенного АТ-нуклеотидами. Участок, с которого вилка репликации начинает двигаться в противоположных направлениях, называется началом двунаправленной репликации (OBR — origin bidirectional replication).
Вспомогательные элементы (Aux) связывают димеры Т-антигена (Аux-1) и фактор транскрипции Sp1 (Аuх-2). Расстояние между этими элементами и их ориентация играют важную роль в процессе инициации репликации. Схема ориджина вируса SV40 представлена на рис. 4.
У эукариот гомологами ориджинов репликации являются автономно реплицирующиеся последовательности, или ARS (autonomously replicating sequences), открытые в 1980 г. Р. Дэйвисом и Дж. Карбоном.

Рис. 4. Схема ориджина вируса SV40.
У дрожжей Saccharomyces cerevisiae особые последовательности, способные обеспечивать репликацию фрагментов ДНК в дрожжевой клетке были выделены раньше, чем у других эукариот. Позднее такие последовательности были найдены и у многих других организмов. У S.cerevisiae АRS занимает 100—200пн и содержит специфическую консенсусную последовательность (АСS — ARS consensus sequence), размером в 11пн, необходимую для связывания с белком-инициатором, а также дополнительные элементы (В-элементы), усиливающие функцию ориджина. Например, АRS1 — первый подробно охарактеризованный ориджин — содержит три таких элемента — В1, В2, ВЗ. Последовательности АCS и В1 занимают приблизительно 50пн и представляют собой наименьшую функциональную область любого ориджина, которая требуется для связывания с белком-инициатором.
Элемент В2 обычно содержит генетически охарактеризованный участок DUE. Вспомогательный элемент ВЗ связывает фактор транскрипции Abf-1. Общая длина ARS-элемента составляет 100-200пн. Строение ориждина S.cerevisiae представлено на рис. 5.

Рис. 5. Схема ориджина Saccharomyces cereiseae
У другого вида дрожжей, Shizosaccharomyces pombe, ориджины состоят по крайней мере из одной ARS, которая значительно длиннее, чем у S. cerevisiae. В некоторых случаях несколько ARS-элементов формируют зону инициации репликации. (Рис. 6.)

Рис. 6. Схема ориджина Shizosaccharomyces pombe
У млекопитающих ориджины детально не охарактеризованы, некоторые из них располагаются в межгенных промежутках, имеют сайты связывания для транскрипционных факторов, часто содержат только районы инициации двунаправленной репликации — OBR.
2.4. Скорость репликации
Скорость репликации генома регулируется в основном частотой инициирующих событий. Так, у Е. соli скорость копирования в каждой репликативной вилке постоянна и равна примерно 1500пн в секунду: следовательно, полный геном длиной 4·10 6 пн реплицируется примерно за 40 мин. Если хромосома реплицируется быстрее, это значит, что увеличивается частота актов инициации в той же самой точке начала репликации при прежней скорости копирования. Клетки Е. соli делятся каждые 20 мин; это означает, что репликация ДНК инициируется в хромосомах, еще не закончивших предыдущий раунд репликации. Скорость движения репликативной вилки в эукариотических клетках значительно меньше (10-100пн в секунду), но завершение репликации хромосомы в разумное время обеспечивается одновременной инициацией во множестве точек. Итак, скорость репликации хромосом контролируется числом и расположением точек начала репликации. Например, в ранних эмбрионах дрозофилы репликация отдельной хромосомы осуществляется каждые 3 мин, благодаря почти одновременной инициации событий в точках, отстоящих друг от друга на 7000-8000пн. В тоже время известно, что у дрозофилы в ходе раннего эмбрионального развития, как скорость репликации, так и размеры и число репликонов тканеспецифичны. В культуре же соматических клеток той же дрозофилы скорость удвоения хромосом значительно более медленная, так как репликация начинается в гораздо меньшем числе точек, находящихся друг от друга на расстоянии 40000пн, при этом продолжительность S-фазы составляет 600 мин. Следовательно, при фиксированной скорости синтеза ДНК множественная инициация повышает скорость процесса репликации в целом и таким образом уменьшает время, необходимое для удвоения всего набора хромосом. Данные о числе репликонов и скорости репликации приведены в табл.1.
Различия в продолжительности S-фазы найдены и у других организмов. Например, у тритона S-фаза длится 1 ч в ядрах бластулы и 200 ч в предмейотической S-фазе сперматоцитов. Вероятно, длительность S-фазы определяется не скоростью синтеза ДНК, а числом задействованных ориджинов репликации. В ДНК клеток нейрулы тритона они находятся на расстоянии около 40 мкм друг от друга, а в соматических клетках — около 100 мкм.
Число и длина репликонов у разных организмов.


В соответствии с современными представлениями репликоны у эукариот распределены в геноме не случайно, они расположены группами (replicon foci). В этих группах, или фокусах, собираются ферменты репликации, которые удлиняют вилки репликации одновременно 10-100 соседних репликонов длиной примерно по 100тпн каждый. Репликация в них завершается за 45–60 мин. Кроме этого существуют очень длинные репликоны (более 1000тпн) — столь большие, что репликация в них продолжается по нескольку часов.
Репликация
1. Когда происходит репликация? – В синтетической фазе интерфазы, задолго до деления клетки. Период между репликацией и профазой митоза называется постсинтетическая фаза интерфазы, в нем клетка продолжает расти и проверяет, правильно ли произошло удвоение.
2. Если до удвоения было 46 хромосом, то сколько будет после удвоения? – Количество хромосом при удвоении ДНК не изменяется. До удвоения у человека 46 одинарных хромосом (состоящих из одной двойной цепочки ДНК), а после удвоения – 46 двойных хромосом (состоящих из двух одинаковых двойных цепочек ДНК, соединенных между собой в центромере).
3. Зачем нужна репликация? – Чтобы во время митоза каждая дочерняя клетка могла получить свою копию ДНК. При митозе каждая из 46 двойных хромосом делится на две одинарные; получается два набора по 46 одинарных хромосом; эти два набора расходятся в две дочерние клетки.
Три принципа строения ДНК
Полуконсервативность – каждая дочерняя ДНК содержит одну цепочку из материнской ДНК и одну новосинтезированную.
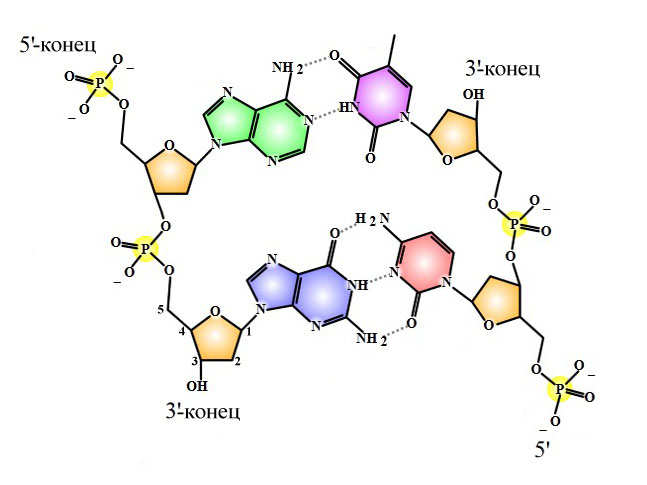
Комплементарность – АТ/ЦГ. Напротив аденина одной цепи ДНК всегда стоит тимин другой цепи ДНК, напротив цитозина всегда стоит гуанин.
Антипараллельность – цепочки ДНК лежат друг к другу противоположными концами. Эти концы не изучают в школе, поэтому чуть подробнее (и далее – в дебри).
Мономером ДНК является нуклеотид, центральной частью нуклеотида – дезоксирибоза. У неё 5 атомов углерода (на ближайшем рисунке у левой нижней дезоксирибозы атомы пронумерованы). Смотрим: к первому атому углерода присоединяется азотистое основание, к пятому – фосфорная кислота данного нуклеотида, третий атом готов присоединить фосфорную кислоту следующего нуклеотида. Таким образом, у любой цепочки ДНК есть два конца:
Правило антипараллельности состоит в том, что на одном конце двойной цепи ДНК (например, на верхнем конце ближайшего рисунка) одна цепь имеет 5′-конец, а другая 3′-конец. Для процесса репликации важно, что ДНК-полимераза может удлинять только 3′-конец. Цепочка ДНК может расти только своим 3′-концом.
На этом рисунке процесс удвоения ДНК идет снизу вверх. Видно, что левая цепочка растет в том же направлении, а правая – в противоположном.
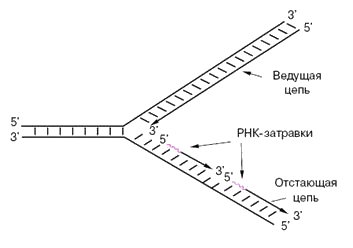
На следующем рисунке вверхняя новая цепочка («ведущая цепь») удлиняется в том же направлении, в котором происходит удвоение. Нижняя новая цепочка («отстающая цепь») не может удлиняться в том же направлении, потому что там у нее 5′-конец, который, как мы помним, не растёт. Поэтому нижняя цепочка растет с помощью коротких (100-200 нуклеотидов) фрагментов Оказаки, каждый из которых растет в 3′-направлении. Каждый фрагмент Оказаки растет от 3′-конца праймера («РНК-затравки», на рисунке праймеры красные).
Ферменты репликации
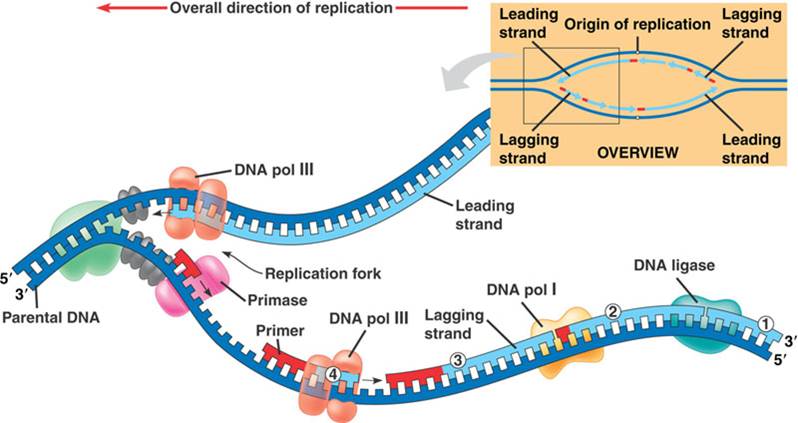
Overall direction of replication – направление, в котором происходит удвоение ДНК.
Parental DNA – старая (материнская) ДНК.
Зеленое облако рядом с надписью «Parental DNA» – фермент хеликаза, который разрывает водородные связи между азотистыми основаниями старой (материнской) цепочки ДНК.
Серые овальчики на только что оторванных друг от друга цепочках ДНК – дестабилизирующие белки, которые не дают цепочкам ДНК соединиться.
DNA pol III – ДНК-полимераза, которая присоединяет новые нуклеотиды к 3′-концу верхней (лидирующей, синтезирующейся неприрывно) цепочки ДНК (Leading strand).
Primase – фермент праймаза, которая делает праймер (красную деталь от Лего). Теперь считаем праймеры слева направо:
На суперкартине не обозначен фермент топоизомераза, но дальше а тестиках он будет фигурировать, так что скажем и про него пару слов. Вот вам веревка, состоящая из трех больших жил. Если три товарища возьмутся за эти три жилы и начнут тянуть их в три разные стороны, то очень скоро веревка перестанет расплетаться и завьется в тугие петли. С ДНК, которая представляет собой двухжильную веревку, могло бы произойти то же самое, если бы не топоизомераза. 
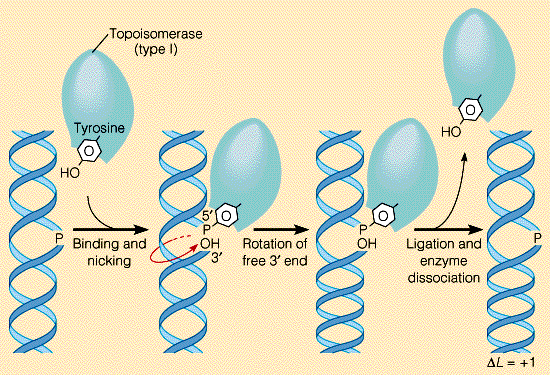
Топоизомереза разрезает одну из двух нитей ДНК, после чего (второй рисунок, красная стрелка) ДНК проворачивается вокруг одной из своих цепей, так что тугие петли не образуются (топологический стресс снижается).
Концевая недорепликация
Из суперкартины с ферментами репликации понятно, что на месте, оставшемся после удаления праймера, ДНК-полимераза достраивает следующий по счету фрагмент Оказаки. (Правда понятно? Если что, фрагменты Оказаки на суперкартине обозначены цифрами в кружочках.) Когда репликация на суперкартине дойдет до своего логического (левого) конца, то у последнего (крайнего левого) фрагмента Оказаки не будет «следующего», поэтому некому будет достроить ДНК на пустом месте, получившемся после удаления праймера.
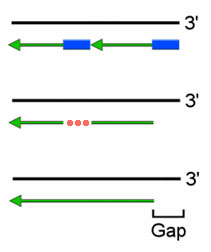
Вот вам еще рисунок. Черная цепочка ДНК – старая, материнская. Удвоение ДНК, в отличие от суперкартины, происходит слева направо. Поскольку у новой (зеленой) ДНК справа 5′-конец, то она является отстающей и удлиняется отдельными фрагметами (Оказаки). Каждый фрагмент Оказаки растет от 3′-конца своего праймера (синего прямоугольника). Праймеры, как мы помним, удаляются ДНК-полимеразой, которая на этом месте достраивает следующий фрагмент Оказаки (этот процесс обозначен красным многоточием). На конце хромосомы некому заделать этот участок, так как нету следующего фрагмента Оказаки, там уже пустое место (Gap). Таким образом, после каждой репликации у дочерних хромосом укорачиваются оба 5′-конца (концевая недорепликация).
Чтобы концевая недорепликация не приводила к печальным последствиям, на концах хромосом имеются участки, не несущие наследственной информации – теломеры. Их укорочение не приносит вреда; у человека они рассчитаны примерно на 60 репликаций. Больше 60 раз (число Хейфлика) клетки человека поделиться не могут, поскольку концевая недорепликация начинает затрагивать гены.
Стволовые клетки (в коже, красном костном мозге, семенниках) должны делиться гораздо больше, чем 60 раз. Поэтому в них функционирует фермент теломераза, который после каждой репликации удлиняет теломеры. Теломераза удлиняет выступающий 3′-конец ДНК, так что он увеличивается до размера фрагмента Оказаки. После этого праймаза синтезирует на нем праймер, и ДНК-полимераза удлиняет недореплицированный 5′-конец ДНК.
Тестики
2. Соотнесите функции ферментов, участвующих в репликации прокариот, с их названиями.
| Ферменты | Функции |
| 1) ДНК геликаза | а) синтез РНК-праймера на отрезке прерывной репликации |
| 2) Праймаза | б) раскрутка двуцепочечной ДНК |
| 3) ДНК полимераза I 3´→5´-нуклеазная активность | в) удаление праймера РНК |
| 4) ДНК полимераза I 5´→3´-нуклеазная активность | г) сшивание пробелов между фрагментами Оказаки |
| 5) ДНК лигаза | д) удаление неспаренных нуклеотидов |
| 6) Топоизомераза | е) снижение топологического стресса при расщеплении двуцепочечной ДНК |
3. Во время репликации в эукариотических клетках удаление праймеров
А) осуществляется ферментом только с ДНК-азной активностью
Б) образует фрагменты Оказаки
В) происходит только в отстающих цепях
Г) происходит только в ядре
4. Если Вы проэкстрагируете ДНК бактериофага fX174, вы обнаружите, что в его составе находится 25% A, 33% T, 24% G, и 18% C. Как Вы могли бы обьяснить эти результаты?
А) Результаты эксперимента неправильные; где-то произошла ошибка.
Б) Можно было бы допустить, что процентное содержание A приблизительно равно таковому T, что также справедливо для C и G. Следовательно, правило Чаргаффа не нарушается, ДНК является двуцепочечной и реплицируется полуконсервативно.
В) Поскольку процентные соотношения A и T и, соответственно, C и G различные, ДНК представляет собой одну цепь; она реплицируется при помощи особенного фермента, следующего особенному механизму репликации с одной цепью в качестве матрицы.
Г) Поскольку ни A не равно T, и ни G не равно C, то ДНК должна быть одноцепочечной, она реплицируется путем синтеза комплементарной цепи и использованием этой двуцепочечной формы как матрицы.
5. Диаграмма относится к репликации двуцепочечной ДНК. Для каждого из квадратов I, II, III выберите один фермент, который функционирует на этом участке.
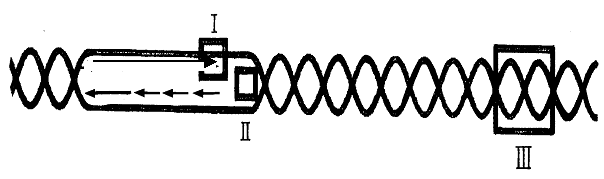
А) Теломераза
Б) ДНК-топоизомераза
В) ДНК-полимераза
Г) ДНК-геликаза
Д) ДНК-лигаза
6. Культура бактерий из среды с легким изотопом азота (N-14) перенесли в среду, содержащую тяжелый изотоп (N-15) на время, соответствующее одному делению, а затем вернули в среду с легким изотопом азота. Анализ состава ДНК бактерий после периода, соответствующего двум репликациям, показал:
| Варианты ответа | ДНК | ||
| легкая | средняя | тяжелая | |
| А | 3/4 | 1/4 | — |
| Б | 1/4 | 3/4 | — |
| В | — | 1/2 | 1/2 |
| Г | 1/2 | 1/2 | — |
7. Одно редкое генетическим заболевание характеризуется иммунодефицитом, отставанием в умственном и физическом развитии и микроцефалией. Предположим, что в экстракте ДНК пациента с этим синдромом вы обнаружили почти одинаковые количества длинных и очень коротких отрезков ДНК. Какой фермент у этого пациента наиболее вероятно отсутствует/дефектный?
А) ДНК-лигаза
Б) Топоизомераза
В) ДНК-полимераза
Г) Геликаза
8. Молекула ДНК, представляет собой двойную спираль, содержащую четыре различных типа азотистых оснований. Какое из следующих утверждений в отношении как репликации, так и химического строения ДНК, является правильным?
A) Последовательности оснований двух цепей одни и те же.
B) В двойной цепи ДНК содержание пуринов равно содержанию пиримидинов.
C) Обе цепи синтезируются в направлении 5’→3’ непрерывно.
D) Присоединение первого основания вновь синтезируемой нуклеиновой кислоты катализируется ДНК-полимеразой.
E) Активность ДНК-полимеразы по исправлению ошибок осуществляется в направлении 5’→3’.
9. Большинство ДНК-полимераз обладает также активностью:
А) лигазной;
Б) эндонуклеазной;
В) 5′-экзонуклеазной;
Г) 3′-экзонуклеазной.
Линейная одноцепочечная ДНК (ssDNA) длиной 6000 нуклеотидов была гибридизована с короткой (300 нуклеотидов) комплементарной одноцеаочечной ДНК, меченой радиоактивными нуклеотидами (a). Эта гибридизованная ДНК была обработана одним из трех способов: ДНК-хеликазой, кипячением без геликазы или прокипяченной геликазой. Затем образцы ДНК были подвергнуты электрофорезу в агарозном геле. На рисунке b показаны полосы ДНК, которые можно было обнаружить в геле при помощи авторадиографии. (Предположено, что необходимая для этой энзиматической реакции энергия АТФ была предоставлена во время обработки ДНК-хеликазой). 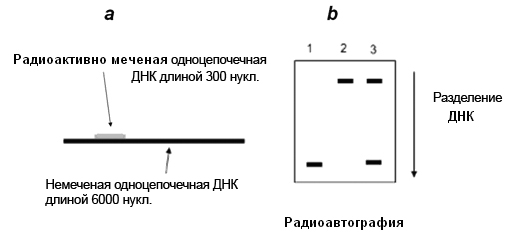
Какое из следующих утверждений относительно этого эксперимента является правильным?
А) Полоса, появляющаяся в верхней части геля, является только ssДНК, величиной 6,3 kb.
Б) Полоса, появляющаяся в нижней части геля, это меченная 300bp ДНК.
В) Если гибридизованную ДНК обработать только ДНК хеликазой и довести реакцию до конца, расположение полос выглядит так, как изображено на дорожке 3 на рисунке b.
Г) Если гибридизованную ДНК обработать только кипячением без обработки хеликазой, расположение полос выглядит как изображено на дорожке 2 на рисунке b.
Д) Если гибридизованную ДНК обработать только прокипяченной хеликазой, расположение полос выглядит как изображено на дорожке 1 на рисунке b.